Lexicon of Evolutionary Genetics
and Related Topics
Compiled by
C. Patrick Doncaster
Compiled by
C. Patrick Doncaster
ADAPTATION. (1) A set of characteristics of an organism which have evolved as a consequence of natural selection in its evolutionary past, and which result in a close match with features of the environment or constrain the organisms to life in a narrow range of environments. The present match between organisms and their environment, and the constraints on this match, have been determined by evolutionary forces acting on ancestors; the heritable characteristics of an organism are consequences of the past and not anticipation of the present or future. The function of an adaptation is not always equivalent to its effect: see TERRITORIAL BEHAVIOUR. (2) Changes in the form or behaviour of an organism during its life as a response to environmental stimuli, e.g. the formation of sun and shade leaves on the same tree and the acquisition of cold tolerance as a result of prior experience of low temperatures. (3) Changes in the excitability of a sense organ as a result of continuous stimulation.
ADAPTIVE LANDSCAPE. A visualisation of the combined effect on average fitness of variation at several loci. A simple landscape map plots fitness contours on two orthogonal axes, representing frequency of allele A and frequency of allele B. If heterozygous advantage at one locus results in MEAN FITNESS being highest when A is present at a frequency of 50%, but heterozygous disadvantage at the other locus results in mean fitness being lowest when B occurs at a frequency of 50%, then the contour map has two adaptive peaks, at {50,0} and {50,100}. If a population has relative frequencies of alleles A and B which locate it on one of the down-slopes, say at {40,20}, natural selection will act on the population to alter the frequencies of A and B such that mean fitness increases. It will therefore move towards the nearest peak, at {50,0}, even if this is not the highest peak in the adaptive zone. Peak shifts might occur through genetic drift, acting on a reduced population and capable of moving it in any direction, even into a trough, from which it can then climb a neighbouring peak through natural selection. The shape and distribution of the contours is determined by the natural environment, which is typically not constant from generation to generation. Thus peaks are constantly changing position and height, and populations are always moving towards the nearest without ever arriving, as expressed by the RED QUEEN'S HYPOTHESIS. It has been suggested that quickly evolving parasites could keep landscapes of adaptation topographically unstable, thus perpetuating the exploration of different peaks and thereby fostering macroevolutionary changes, compared to the microevolution of hill climbing.
ADAPTIVE ZONES. Clusters or constellations of adaptive peaks in an adaptive landscape, separated from each other by major adaptive troughs, yielding natural ecological groupings of species.
ADENOSINE TRIPHOSPHATE. (ATP). High-energy phosphoric nucleotide of the nucleoside adensosine which functions as the principal energy carrying compound in the cells of all living organisms. Its hydrolysis to adenosine diphosphate (ADP) and inorganic phosphate is accompanied by the release of a large amount of free energy, which is used to drive many metabolic functions.
ALLELE. One of two or more alternative forms of a gene, distinguishable from other forms or alleles of the same gene. For example, a species of flowering plant contains two alleles a1 and a2 which control the colour of the seed coats; a single recessive gene controls the trait of albinoism with two alleles a (albino) and A (normal). Two alleles on one locus produce three diploid genotypes: AA, Aa, aa. Addition of a second locus with two alleles B and b makes nine genotypes possible (AABB, AABb, AaBB, ...). In general, where each locus contains just two alleles, the number of possible genotypes in a population containing n of the loci is 3n. In fact the number of alleles is usually higher, and so the general case is: the total number of genotypes that can be put together from all the n genotypes is m1 x m2 x m3 x ... x mn, where m1 is the number of genotypes possible at the first locus, m2 at the second, and so on. In humans, at least 10,000 loci exist, many of which carry numerous alleles. So the total number of diploid genotypes is astronomical. The commonness of an allele in a population is termed the `allele frequency.'
ALLOPARENT. An individual, other than the genetic parent, that provides care for conspecific young. Alloparents may acquire selective advantages associated with increased inclusive fitness, parental experience, reciprocal altruism, and exploitation of fostered young [Riedman, M.L. 1982. Quart. Rev. Biol., 57:405].
ALTRUISM. Acting to increase the lifetime number of offspring of another individual at a cost to the performer's own chances of survival and reproduction (its individual FITNESS). See KIN SELECTION.
ANAGENESIS. Evolutionary change with time (descent with modification) within a single lineage (see also CLADOGENESIS).
ANTIGEN. Any substance that the body regards as foreign and that therefore illicits an immune response. A typical immune response would be production of lymphocyte white blood cells composed of antibody proteins which react with the antigens to render them harmless. Antibody-antigen reactions are highly specific.
APOMORPH. Evolutionarily advanced (derived) character state. For example the long neck of the giraffe is apomorphic, the shorter neck of its ancestor is plesiomorphic.
ASEXUAL REPRODUCTION. Reproduction that does not involve meiosis or the production or fusion of gametes. In plants it consists of vegetative reproduction, apospory and apogamy. Asexual reproduction occurs by fission in many protozoan animals but also in some multicellular ones (e.g. corals); and by budding (gemmation) in many liverworts, mosses, coelenterates and ascidians. The advantages of asexual over sexual reproduction are that a mate is not required, and it is quicker and less energetically costly; the principal disadvantage is that individuals do not exchange genetic material and so their offspring lack the genetic diversity to buffer against environmental change. Although many sexual organisms can also reproduce asexually when rapid generation of young is advantageous (e.g. parthenogenesis by aphids in spring and summer), few organisms are entirely asexual. Indeed, the diverse and substantial advantages of SEXUAL REPRODUCTION raise the question of how any organism can survive through evolutionary time without sex. Asexuality in a taxon, such as the bdelloid rotifers, may be due to the frequent production by a core sexual species of asexual mutants. These new lineages initially flourish at the expense of sexual forms which, in terms of individual fitness, have too high a degree of genetic diversity; but each go extinct after a few hundred years through their inability to adapt to changing environments. [See Hurst, L.D. et al. 1992. TREE, 7:144].
AUTOSOME. Any chromosome in the cell nucleus other than a SEX CHROMOSOME.
BACK CROSS. A cross-fertilisation made to identify hidden recessive alleles. An organism possessing a dominant characteristic can be shown to be homozygous for the character or heterozygous, by crossing it with one containing the recessive characteristic. If all the offspring have the dominant characteristic, then it is homozygous, if only half have it then it is heterozygous.
CENTRAL DOGMA. In molecular biology, the dogma that nucleic acids act as templates for the synthesis of proteins, but never the reverse. More generally, the dogma that genes exert an influence over the form of a body, but the form of a body is never translated back into genetic code: acquired characteristics are not inherited. Accredited to F. Crick.
CHAOS. Fluctuations that have no fixed periodicity. For example gradually increasing the size of a parameter for `boom-or-bustness' in a population (a combination of strength of density-dependence and net reproductive rate) causes changes in the type of equilibrium population size. At low parameter values the population follows damped oscillations which approach a single equilibrium with time. When the parameter is above a critical level the population starts to oscillate between two stable sizes (i.e. to become cyclic in time within stable limits); above a higher critical level the two equilibria each bifurcate again to give four stable sizes (a cycle with two periods); higher again it bifurcates again to give eight; and then as the system is being pushed very hard by ever higher parameter values it breaks down into fluctuations with no periodicity. High reproductive rates, overcompensating density-dependence and time-lags are capable of provoking all types of fluctuations in population density, including chaotic, without invoking any extrinsic cause. [See Grenfell, B.T. et al. 1992. Nature, 355:823]
CHARACTER DISPLACEMENT. Phenomenon whereby competition between closely related species causes them to become more different in regions where their ranges overlap than in regions where they do not. The 14 extant species of Galapagos finches are a classic example of character displacement associated with geographic speciation. They evolved from a single species which divided many times over on the Galapagos islands through the process of MULTIPLE INVASION. As newly formed species accumulated they faced the problem of interference through competition and hybridisation. For example, three seed-eating species have different depths of bill where they are sympatric, and thus avoid competition by feeding on different kinds of seeds; bill depth only overlaps where they are allopatric.
CHROMOSOME. Complex structure found in the nucleus of a eucaryotic cell, composed of nucleic acids and proteins, and bearing part of the genetic information (genes) of the cell. Humans have 46 chromosomes (analogy: 46 filing cabinet drawers each containing pages, the genes, of DNA code, A-C-G-T..., written on them). The chromosomes are usually present in all cells in the body, even though only a minority of them will be active in any one cell.
CLADE. The set of all species descended from a single ancestral species.
CLADISTICS (PHYLOGENETIC SYSTEMATICS). A systematic method whereby taxa are hierarchically clustered according to inferred relative recency of common ancestry, based upon their shared derived character states. It is based on the assumption that two new species are formed suddenly, by splitting from a common ancestor, and not by gradual evolutionary change. It has been used to argue that the major branching patterns of cladograms correspond to large-scale evolutionary events that cannot be explained by orthodox neo-Darwinism.
CLADOGENESIS. Branching of an evolutionary lineage, resulting from the splitting of one species into two or more. (See also ANAGENESIS; SPECIATION).
CLINE. Continuous gradation of form or gene differences in a population of a species, correlated with its geographical or ecological distribution. Clines usually take one of two forms: North/South, for example the spotting pattern of the Large Heath butterfly (Coenonympha tullia) in the British Isles; or altitudinal, for example the spotting pattern of the Corsican Heath butterfly (Coenonympha corinna) in Corsica.
COEFFICIENT OF RELATEDNESS. The fraction of genes identical by descent between two individuals. Identical twins have a relatedness of 1, full sibs of 1/2, half sibs of 1/4, cousins of 1/8. Thus an individual helping a full sib to have two offspring has the same effect on the gene frequency as if it were able to have one offspring itself. A useful measure for applying to HAMILTON'S RULE and INCLUSIVE FITNESS theory is r defined as the extent to which a potential actor helping a potential recipient is like the actor helping itself. [See p. 13 of Krebs & Davies, 1991 for different methods of measuring relatedness].
COMPARATIVE METHOD. A statistical approach to analysing broad trends in evolution, and the general relationship between social organisation and ecology. It generates hypotheses which can be used as predictions for other groups of animals, and can also be used to test hypotheses not amenable to experimentation. The method has been employed for example to discriminate between competing hypotheses for the evolution in sexual dimorphism between primate males (large) and females (small). If sexual dimorphism is an evolutionary solution to competition between the sexes for food, then dimorphism is predicted to be strongest in monogamous species where the sexes associate and feed together. But if it has evolved through sexual selection, with larger males out-performing smaller males in competition for females, then dimorphism should be greater in polygamous species where a large male could potentially monopolise several females. The comparative data show trends which support the second hypothesis and not the first. Such statistical analyses must be based on independent evolutionary events, and species are not necessarily appropriate data points. Ideally, comparisons should be made between all ancestor-descendent pairs on a known phylogenetic tree. Where ancestral character states are not known they can be guessed as the mid-point between two descendent species. [See p.36 of Krebs & Davies, 1993].
COMPETITION. Interaction between individuals brought about by a shared requirement for a resource in limited supply, and leading to a reduction in one or more components of fitness (survivorship, growth, reproduction). Competition produces much of the variation in fitness on which natural selection acts.
CONVERGENT EVOLUTION. The development of similar characteristics in organisms that are not closely related, as each adapts to a similar way of life. Sharks (fish), dolphins (mammals), and ichthyosaurs (extinct reptiles) are examples of convergence in the aquatic habitat.
COUNTERVAILING SELECTION. Selection that supports different genotypes in different fitness components. Individual fitness components can thus have high heritabilities while overall fitness has a low heritability. For example, genotypes favouring reproductive performance in female red deer tend to be those that survive poorly as juveniles [Pemberton et al. 1991. Evolution, 45:93].
CROSSING OVER. Exchange of parts between homologous chromosomes through a process of breakage and reunion.
CYTOPLASMIC INHERITANCE. Non-Mendelian (extra-chromosomal) inheritance via genes in cytoplasmic organelles. Examples of such organelles are viruses, mitochondria and plastids (e.g. chloroplasts). These all have their own genetic material, which is open to mutations that do not follow Mendel's rules for chromosomal genes. Although both male and female parents contribute equally to the zygote in terms of chromosomal genes, it is usually only the female parent which contributes the initial cytoplasm and organelles of the zygote. Zygotic development therefore usually begins within a maternal milieu, so that maternal cytoplasm directly affects zygotic development. (See also SEX).
CYTOPLASM. The part of the cell that is enclosed by the plasma membrane, but excluding the nucleus.
DEGENERACY. The situation where more than one sequence of bases in the DNA molecule codes for the same (amino acid) product.
DEME. A spatially discrete local population within which breeding is completely random: all possible male and female pairings within a deme have an equal chance of forming, for one breeding season at least. A deme is hence the largest population unit that can be analysed by the simpler models of population genetics.
DISPERSAL. The tendency of an individual to move away from its birth area
(natal dispersal) or breeding area (breeding dispersal), in search of improved
feeding or mating opportunities. Philopatry is the opposite tendency, to
return or stay in its home area, inheriting a share in its resources and
avoiding the risks associated with seeking a new area. In birds and mammals one
sex usually disperses more than the other, with the result that close
inbreeding is avoided. The direction of bias is associated with the type of
MATING SYSTEM. In general, dispersal tends to be favoured by the sex that is
limited and philopatry by the sex that is limiting; a male bias in dispersal is
usually associated with a mate-defence mating system (males limited by
females), and a female bias with a resource defence system (females limited by
resources). [See Greenwood, P.J. 1980. Anim. Behav., 28:1140; Greenwood,
P.J. 1983. In: The Ecology of Animal Movement. Clarendon, Oxford.] Thus,
the majority of mammals exhibit mate-defence POLYGYNY (probably in response to
resource distribution and quantity, or predation threat), and males disperse in
the process of competing for a limited supply of females; females are
philopatric, defending the food resources required for gestation and weaning of
offspring. The majority of birds exhibit resource-defence MONOGAMY (probably in
response to costs of incubation and brooding), and males compete for a limited
supply of resources with which to attract females, thereby gaining home
advantage from philopatry; females disperse in the process of choosing between
the available resources of different males. Ethiopian wolves show female-biased
dispersal, associated with reproductive suppression by the pack's
 female (to ensure cohesion of a sufficiently large pack to defend a very
limited supply of good quality habitat patches against other packs) which
results in subordinate females only gaining breeding opportunities away from
the natal range; males never disperse but remain as philopatric helpers, in the
absence of unoccupied quality habitat to settle elsewhere, and without a
mechanism for joining other packs. Communally breeding Florida scrub jays
exhibit a similar system, where habitat saturation again limits breeding
opportunities. For wild dogs the stable patrilinear groups and monogamous
breeding is regulated through reproductive suppression by the female,
which functions to guarantee sufficient cohesion of the group for pack hunting
of prey species that are abundant but too large to be predated by individuals.
The inclusive fitness of males may be improved by philopatry, as they can help
the female rear a very large litter of 10-20 pups; they may also gain
mating opportunities with the female (polyandry). Male philopatry in
primates is associated not with resource defence, but with the tendency for
males to cooperate in intra-sexual competition (forming alliances against males
in neighbouring groups). See also HOMOGAMETIC SEX.
female (to ensure cohesion of a sufficiently large pack to defend a very
limited supply of good quality habitat patches against other packs) which
results in subordinate females only gaining breeding opportunities away from
the natal range; males never disperse but remain as philopatric helpers, in the
absence of unoccupied quality habitat to settle elsewhere, and without a
mechanism for joining other packs. Communally breeding Florida scrub jays
exhibit a similar system, where habitat saturation again limits breeding
opportunities. For wild dogs the stable patrilinear groups and monogamous
breeding is regulated through reproductive suppression by the female,
which functions to guarantee sufficient cohesion of the group for pack hunting
of prey species that are abundant but too large to be predated by individuals.
The inclusive fitness of males may be improved by philopatry, as they can help
the female rear a very large litter of 10-20 pups; they may also gain
mating opportunities with the female (polyandry). Male philopatry in
primates is associated not with resource defence, but with the tendency for
males to cooperate in intra-sexual competition (forming alliances against males
in neighbouring groups). See also HOMOGAMETIC SEX.
DNA (DEOXYRIBONUCLEIC ACID). The fundamental hereditary material of all living organisms. DNA is composed of a double-stranded helix of sugar phosphate held together by pairs of nucleotide bases in linear sequence. These nitrogenous bases are thymine, cytosine, guanine and adenine (C-T-G-A), and the four bases in the two strands are always paired in such a way that A lies opposite T and G lies opposite C. DNA embodies the genetic code. Because of the complementary base-pairing the two strands of DNA can serve as original and template for the production of a new complementary partner strand. DNA therefore carries information by means of the linear sequence of its nucleotides, each of which can be considered as a letter in a simple four-letter alphabet that is used to write out biological messages in a linear `ticker-tape' form. Since the number of different possible sequences of a DNA chain that is n nucleotides long is 4n, the amount of biological variety that can be generated using even a modest length of DNA is enormous. A single human cell comprises 1.5 m of DNA (5 x 109 nucleotide base pairs), organised into the 46 chromosomes that carry our genes. A single gene comprises about 1000 base pairs.
DNA FINGERPRINTING. The variable-number-of-tandem-repeats (VNTR) locus (one of the hypervariable loci) contains many tandem repeats of short segments and, due presumably to unequal crossing over, many alleles are generated at this locus (any one person has, of course, only two of the many possible alleles). Where a single probe can locate several such loci, each individual will produce many bands on a Southern blot, with most people having unique patterns.
EPISTATIC EFFECT. The production in an organism of a character determined by genes at two or more loci, different from the characters coded individually at each locus.
EUKARYOTES. Organisms with cells having true nuclei (i.e. from the green algae upwards). See PROKARYOTES.
EVOLUTION. Any changes in the genetic composition (gene frequencies) of a population during successive generations. The four agents of evolution are: MUTATION PRESSURE, GENETIC DRIFT, GENE FLOW, NATURAL SELECTION.
EVOLUTIONARILY STABLE STRATEGY (ESS). A strategy which, if adopted by most members of a population, cannot be bettered by any other strategy. Because it resists invasion by mutant strategies, it therefore becomes established by natural selection. For example, the optimum time for a male dungfly to wait for a female on a cowpat depends on what all the other males are doing, so the pay-offs for different waiting times are frequency dependent. The ESS is to play a randomly chosen waiting time (to be unpredictable); if this is heritable and individuals reproduce in proportion to their pay-offs, then the ESS will spread in the population. The ESS therefore predicts that waiting times will be variable, either within or between individuals, and that pay-offs will be equal as a result of FREQUENCY-DEPENDENT SELECTION. A mixed ESS can result from frequency-dependent selection driving the frequencies of two possible strategies (e.g. Hawks and Doves) in the population so that they each enjoy equal average pay-offs. If the population moves away from the equilibrium towards more of one strategy then the other strategy does better, driving it back to the equilibrium mixture. Where there is more than one ESS for a given game, populations should evolve to different behavioural equilibria depending on their initial composition; competition between these groups then leads to the best ESS prevailing over inferior types. Because it is that ESS which bestows on its group the greatest proliferative advantage compared to other ESSs for the same game, the advantage of the best ESS accrues to the group, not to the individual. ESSs at equilibrium are therefore true group adaptations; the ESS that goes to fixation is not necessarily that which serves each individual practitioner's interests best [see Koeslag, J.H. & Koeslag, P.D. 1994 J. theor. Biol., 167:55]. For example, in the game of reproduction, with two competing strategies of asex and bi-gender sex, a panmictic population has an ESS in asex while a koinophilic population has an ESS in bi-gender sex. But koinophila is itself an ESS with respect to panmixis, even when the incidence of new alleles includes an unusually high fraction of beneficial mutations.
FISHER'S FUNDAMENTAL THEOREM. The rate of increase in fitness is equal to the additive genetic variance in fitness. This can be expressed by R = h2×S, where R is the amount by which the mean value for a character in the offspring generation differs from that in the parental population: a measure of the rate of evolution, h2 is HERITABILITY, S is the SELECTION DIFFERENTIAL. Strong selection on a character leads to rapidly decreasing variation in that character, S therefore becomes smaller and consequently the rate of evolutionary change is reduced. [See Gustafsson, L. 1986. Am. Nat., 128:761.]
FISHER'S HYPOTHESIS. An elaborate male display may be sexually selected simply because it makes males attractive to females. Thus a female choosing a mate with the trait gains the advantage of sons that are attractive to females. The initial starting point may have been for a trait with utilitarian benefit (e.g. males with longer tails were better flyers), but the positive feedback between female preference and the trait can then take over from the benefits to viability, to the point where the sexual attractiveness of the trait counter-balances the decrease in survival ability it entails. The main competitor to this hypothesis is the HANDICAP HYPOTHESIS which assumes that benefits are to do with general viability (e.g. disease resistance), rather than increased mating success alone.
FISHER'S SEX RATIO THEORY. Selection acting on the individual tends to produce population sex ratios close to 1:1 even where a single male can fertilise the eggs of many females. A female-biased sex ratio, say 1M:2F, is not evolutionarily stable because an average male gets twice as many mates as each female, so a gene which causes parents to bias the sex ratio of their offspring towards males would rapidly spread. Conversely, if males are twice as common as females then daughters will be advantaged, because only one sperm fertilises each egg so only half the males can contribute genes to any individual offspring. Only when the sex ratio is exactly 1:1 will the expected success of a male and a female be equal and the population stable. When sons and daughters cost different amounts to make, the stable evolutionary strategy is for the parent to invest equally in the two sexes rather than to produce equal numbers.
FISSION. (allopatric speciation by). The intrusion of a new barrier divides a species into two isolated populations which then evolve into two species. If the barrier then disappears, the two new daughter species can coexist in the same habitat without interbreeding.
FITNESS. The environment, and competition for survival within it, determines which individuals are fittest to survive and propagate their genes in the next generation. An individual can obtain a direct fitness gain if it can increase its own reproductive success, and an indirect fitness gain through aiding the survival of non-descendent kin, such as siblings. The sum of both these routes to fitness gain is a measure of the individual's INCLUSIVE FITNESS. [See also GENETIC FITNESS].
FIXATION. In population genetics, the complete prevalence of one gene form (allele), resulting in the complete exclusion of another.
FLUCTUATING ASYMMETRY. A population phenomenon defined as random deviation from perfect bilateral symmetry in a morphological trait for which differences between the right and left sides have a mean of zero and are normally distributed. Because both sides of bilateral structures are produced by the same genome, they are genetically programmed to develop in a symmetrical manner. Deviations from symmetry are assumed to be due to stress imposed on the individual during the development of the trait; in other words, the degree of symmetry reveals an individual's ability to canalise development in the face of stress. As a measure of individual viability, FA has the advantage of showing deviations from a known morphological ideal (perfect symmetry). Thus, unlike indices of phenotypic quality such as body size or fecundity, for which it is difficult to specify an optimum (which may vary with environmental conditions), asymmetry seemingly provides an unambiguous and consistently valid measure of maladaptation. FA in a sexual traits may reliably reflect the quality of the possessor, and therefore represent an assessment criterion. As relative levels of asymmetry are much greater in secondary sexual characters than in normal morphological traits, it has been proposed that females use the amount of asymmetry in secondary sexual ornamentation as a cue in mate choice. Secondary sexual characters may be more susceptible to disruption during development because they are subject to directional selection which would select against modifiers that may buffer development (non-sexually selected traits are developmentally canalised by genetic modifiers); and mutations are more likely to have a disruptive effect on characters which are elaborate in design. [See Watson, P.J. et al. 1994. TREE, 9:21].
FOUNDER EFFECT. When a new population is derived from a few immigrants, these founders represent a very small sample of the genetic pool to which they formally belonged; natural selection operating on this restricted variety soon yields gene combinations quite different from those found in the ancestral population, or those of a second small sample of founders.
FREQUENCY-DEPENDENT SELECTION. A mode of selection in which the relative fitnesses of different phenotypes within a population changes in relation to their relative frequencies, such that a particular phenotype has a different fitness when rare to when common. Negative frequency dependent selection (fitness improving with rarity) probably plays an important role in maintaining POLYMORPHISM in populations. Examples include FISHER'S SEX RATIO THEORY (offspring of the rarer sex are always favoured); maintenance of polymorphism in prey (predators tend to overlook rare prey types, giving a selective advantage to rare morphs); the rare male effect (females may prefer to mate with a rarer phenotype, although the adaptive value of this MATE CHOICE is obscure); the two mating strategies of male horned beetles, either short-lived but fast reproducing and carrying large horns for fighting, or long-lived with no horns and infrequent sneaky-matings.
GAME THEORY. A technique for modelling frequency dependent behaviours: where the optimal strategy for one individual depends on how other competitors in the population are behaving. Evolution is expected to lead to an EVOLUTIONARILY STABLE STRATEGY. Developed from economic theory principally by J. Maynard Smith, it is most useful in understanding the evolutionary stability of fighting and displaying.
GAMETE. A reproductive cell (haploid) which unites with another in fertilisation to produce a zygote from which a new individual arises.
GENE. A hypothetical unit of heredity. Genes are arranged in linear fashion on the chromosome, and each segregates as a single unit during meiosis and gives rise to a definable phenotypic trait. The "gene for blue eyes" is not one stretch of DNA solely responsible for causing eyes to be a particular colour, but several sections of DNA making enzymes that control various steps of the chemical reactions by which a pigment is synthesised or not. The genes that produce wrinkled seeds in peas comprise the strands of DNA that make the enzymes that critically affect the seed-wall building process, leading to partial loss of moisture and wrinkling. In most cases, a gene corresponds to a region of the genome that directs the synthesis of a single enzyme, so one gene corresponds with one polypeptide chain.
GENE CLONING. Production of large numbers of a piece of DNA after that piece of DNA is inserted into a vector and taken up by a cell. Cloning occurs as the vector replicates.
GENE FLOW. The injection of new genes by migration. In most cases, a deme (a randomly mating population) is only partially isolated from its neighbours of the same species. Where the rate of immigration is high, gene flow can serve both as a source of new genetic variability and as a prime mover of evolution, on a par with natural selection. If immigrants are genetically very different from their host population, then a small amount of immigration can result in a great deal of evolution. The extreme case of such exchange occurs during hybridisation. The mechanism of gene flow is most obviously migration, but it can also be the movement of gametes, the extinction and recolonisation of entire populations, or the movement of extranuclear segments of DNA, such as mitochondria, plasmids and viruses. [See Slatkin, M. 1985. Ann. Rev. Ecol. Syst., 16:393.]
GENE POOL. All of the hereditary material (genes) in a population.
GENETIC CODE. The set of correspondences between base (nucleotide pair) triplets in DNA and amino acids in protein. These base triplets carry the genetic information for protein synthesis. For example, the triplet C-A-A codes for valine, one of the twenty amino acids found in proteins.
GENETIC DRIFT. The alteration of gene frequencies (evolution) through chance processes alone. For example if an Aa individual of Drosophila is crossed with another Aa, the offspring may not yield the exact 1:2:1 Mendelian ratio. If four individuals were selected at random from the offspring to start a new population, chance dictates that other ratios would obtain some of the time, and an allele could be lost entirely or fixed in a single step. Genetic drift is most likely to be effective in very small populations (< 100 individuals), where reduction of genetic variability lowers the capacity of a population to adapt to changes in the environment, and also tends to reduce the overall fitness of the population. INBREEDING DEPRESSION expresses the same effect as small population size in promoting genetic drift, and genetic drift can also operate through the FOUNDER EFFECT: new populations start from a small number of pioneers, which contained only a small fraction of the alleles found in the source population, and so the new population differs at the beginning from its source.
GENETIC FITNESS. The relative contribution that an individual makes to the gene pool of the next generation. The fittest individuals in a population are those that leave the greatest number of descendants relative to the number of descendants left by other, less fit individuals in the population. Those individuals that leave the greatest proportion of descendants in a population have the greatest influence on the heritable characteristics of that population. Different fitness components include first breeding age, fecundity, survival.
GENETIC LOAD. The relative decrease in the mean fitness of a population due to the presence of genotypes that have less than the highest fitness. Genetic load is also the average number of lethal mutations per individual in a population.
GENETIC MARKERS. Molecular methods for making inferences about descent. These include `allozymes', `restriction fragment length polymorphisms' (RFLPs), `multilocus minisatellite' or `DNA fingerprints', `single locus minisatellites' or `variable number of tandem repeats' (VNTRs), `randomly amplified polymorphic DNA' (RAPDs), `microsatellites,' and `mitochondrial DNA' (mtDNA). Amongst the newer methods, minisatellites can provide a similarity index for populations, microsatellites reveal information about kinship, and mitochondrial DNA determines the maternal contribution to descent. [See Queller, D.C. et al. 1993. TREE, 8:285.]
GENOME. The complete genetic constitution of a eukaryotic organism. The full genome of an individual is borne by a single representative of each of all the chromosome pairs in a nucleus. The human genome consists of 3 billion base pairs of DNA, encoding approximately 30,000 genes (the functions of which are unknown for the vast majority).
GENOMIC IMPRINTING. Genes that change their pattern of expression depending on whether they are maternally or paternally inherited are said to have been imprinted by their parental genome. This difference in gene expression has been held to reflect a conflict between paternal and maternal genes, concerning for example the amount of resource transfer from the mother to the developing embryo (with paternal genes being selected to maximise transference of resources from the mother, and maternal genes selected to reduce transference to avoid reducing the mother's chances of survival). [See Hurst, G.D.D. et al. 1992. TREE, 7:373].
GENOTYPE. An exact description of the genetic constitution of an individual, with respect to a single trait, or set of traits. PHENOTYPE is the product of the genotype interacting with the environment. Some expressions of the genotype are resistant to the effects of the environment (blood type, eye colour), others are more readily influenced (stature, athleticism).
GOOD GENES. These models of sexual selection predict that MATE CHOICE should be based on traits that honestly reflect heritable fitness gains. It has been suggested that FLUCTUATING ASYMMETRIES provide just such information. An ornament or HANDICAP may also indicate `bad' mutations; females consequentially preferring males with the handicap in good condition do better than randomly mating females. This is advantageous only for as long as the handicap remains mutation sensitive; males will be under selective pressure to produce heritable fakes. The commonness of a trait, however, which is the fitness indicator that KOINOPHILIC individuals are attracted to, cannot be heritably faked and is therefore likely to be more enduring than the preference for a handicap or other incidental indicator of fitness.
HAMILTON'S RULE. Helping another to breed is favoured over breeding oneself when the cost to the actor (c) is less than the benefit to the recipient (b), weighted by the COEFFICIENT OF RELATEDNESS between them (r): rb > c. Costs and benefits are both measured in number of offspring. For example if the AA and Aa genotypes are altruistic, in a diploid population with A and a alleles at one locus, what is the condition for A to spread when the recipients are all half-sibs? Since r=1/4, A will spread if b/c>4. This rule is more illuminating and easier to apply to field data than is the notion of INCLUSIVE FITNESS from which it is derived, although its interpretation depends on how r is calculated. It should also be used in the form rb-c>0, in case c or r is negative, and in order to obtain confidence limits for rb-c simply from the sampling variances of b and c [see p. 20 of Krebs & Davies, 1991]. Hamilton's rule has also been extended by R.L. Trivers to cover the theory of parent-offspring conflict. A mother's optimum investment in an offspring is a trade-off between increasing the offspring's fitness by allocating resources to it, and not diminishing her future reproductive success by investing too highly in the current breeding season. The offspring's demand for resources are also influenced by this trade-off, as it too has a genetic interest in future sibs. However, while the mother is identically related to all her progeny, the offspring values investment in itself twice as much as in future full sibs (and 4 times as much as in half-sibs). A conflict of interest thus arises between the mother and her offspring, with the offspring demanding more resources than the mother is willing to give.
HAPLOID. Having a chromosome complement consisting of just one copy of each chromosome; c.f. DIPLOID, consisting of two copies (homologues) of each chromosome. A diploid individual usually arises as a result of the fusion of two haploid GAMETES (egg and sperm). Thus the two homologues in each chromosome pair in a diploid cell are of separate origin, one derived from the female parent and one from the male.
HANDICAP HYPOTHESIS. Some elaborate adornments of male birds such as peacocks, birds of paradise etc may have evolved for use in contests between males, but others seem to have evolved as a result of selection by females for genetic benefits. The handicap hypothesis, first proposed by A. Zahavi, explains how selection for genetic benefits might produce elaborate traits: the long tail of the male peacock is a handicap to day-to-day survival; females prefer long tails precisely because they are a handicap and therefore act as a reliable signal of a male's genetic quality. The adornment demonstrates a male's ability to survive in spite of the handicap it imposes, which means that he must be extra good in other respects. If this tendency to be good at surviving is heritable, then the female selects for `good genes' by selecting to mate only with males whose displays honestly indicate their genetic quality. W.D. Hamilton and M. Zuk have further proposed that sexual displays are reliable indicators of genetic resistance to disease. If males are able to show the full development of their secondary sexual characteristics only when they are free of diseases, then females choosing elaborate displays are acting as diagnostic veterinarians. This version of the handicap hypothesis is attractive because disease is a widespread agent of selection, and diseases have the property of continually evolving new varieties, hence continually presenting new selective hurdles. See also FISHER'S HYPOTHESIS, GOOD GENES and MATE CHOICE.
HARDY-WEINBERG LAW. The allelic (gene) and genotype frequencies in an
infinitely large population remain constant from generation to generation if
mating is at random and there is no selection, migration or mutation. If two
alleles A and a are segregating at a locus, and each has a
frequency of p and q respectively (which can be observed from the
phenotype frequencies if a is not recessive), then the frequencies of
the genotypes AA, Aa, aa are p2,
2pq, q2 respectively. This Hardy-Weinberg ratio is
thus a null expectation on the basis of random mating and no evolutionary
change. For example, immediately following a severe frost, and before breeding
recommences, the phenotype frequencies for spotting in a sample of 1000 moths
are 551 `conspicuous' (corresponding to the assumed AA genotype), 439
`intermediate' (corresponding to the Aa genotype) and 10 `inconspicuous'
(corresponding to the aa genotype). Allele frequencies are then
calculated as proportions of all alleles: p = (2x551+439)/(2x1000) =
0.771, q = 1-p = 0.229. The expected genotype frequencies are
thus p2(AA) = 0.594,
2pq(Aa) = 0.353, and
q2(aa) = 0.053. A chi-squared test shows
that observed and expected frequencies differ significantly
(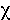 2 = 59.0, with 1 d.f. given by number of phenotypes minus
number of alleles). Thus, after the frost a higher proportion of individuals
possessing the heterozygous condition survive than predicted by the
Hardy-Weinberg formula, and both homozygotes have lower than expected survival:
the moth population can be said to have undergone a small amount of evolution.
If there is no evolutionary change in subsequent generations, and random
mating, then the allele frequencies of 0.771 (A) and 0.229 (a)
will be maintained, establishing new genotype frequencies of 0.594:0.353:0.053.
Where a trait is dominant, allelic frequencies cannot be calculated from the
genotypic classes because the homozygous dominants cannot be differentiated
from the heterozygotes. But for such traits, or for example a disease that is
expressed only in the homozygous recessive state, assuming Hardy-Weinberg
equilibrium makes it possible to calculate the proportion of a population that
is heterozygous. If the genetic disease is present in 1 in 10,000 of the
population, then q2 = 0.0001, so q = 0.01 and thus
p = 0.99. The frequency of the heterozygote is therefore 2pq =
0.02. Thus a recessive gene causing a trait as rare as 1 in 10,000 is carried
in the heterozygous state by 1 in 50 individuals.
2 = 59.0, with 1 d.f. given by number of phenotypes minus
number of alleles). Thus, after the frost a higher proportion of individuals
possessing the heterozygous condition survive than predicted by the
Hardy-Weinberg formula, and both homozygotes have lower than expected survival:
the moth population can be said to have undergone a small amount of evolution.
If there is no evolutionary change in subsequent generations, and random
mating, then the allele frequencies of 0.771 (A) and 0.229 (a)
will be maintained, establishing new genotype frequencies of 0.594:0.353:0.053.
Where a trait is dominant, allelic frequencies cannot be calculated from the
genotypic classes because the homozygous dominants cannot be differentiated
from the heterozygotes. But for such traits, or for example a disease that is
expressed only in the homozygous recessive state, assuming Hardy-Weinberg
equilibrium makes it possible to calculate the proportion of a population that
is heterozygous. If the genetic disease is present in 1 in 10,000 of the
population, then q2 = 0.0001, so q = 0.01 and thus
p = 0.99. The frequency of the heterozygote is therefore 2pq =
0.02. Thus a recessive gene causing a trait as rare as 1 in 10,000 is carried
in the heterozygous state by 1 in 50 individuals.
HELPER. In some species of birds, mammals and fish there are individuals that do not breed themselves, but help others to rear offspring. These helpers are usually, but not invariably, close relatives of the breeders that they help (e.g. young from a previous season). Helping close relatives is a way of promoting genetic representation in the next generation. See HAMILTON'S RULE.
HERITABILITY. Of the total phenotypic variance among individuals in a trait, which is the sum of genetic variance and environmental variance, the proportion that is attributable to differences in genotype: h2 = genetic variance / total phenotypic variance. Natural selection tends to reduce heritability, because it leads to increasing homogeneity in genotype over successive generations as less fit genotypes are eliminated. Heritability is therefore lower for traits that are strongly related to fitness than for other traits.
HETEROZYGOUS. Of a diploid organism having different alleles of a given gene on the pair of homologues carrying that gene. Because a heterozygote has two or more different alleles at a given locus, it does not breed true.
HETEROZYGOUS ADVANTAGE. Situation in which individuals that are heterozygous at a given locus have higher fitness than either of the homozygotes.
HOMOGAMETIC SEX. The sex that is determined by the possession of two similar sex chromosomes (XX). The chromosomes themselves are not determining sex, but the genes they carry are. Genotype determines the type of gonad that then determines the phenotype of the organism through male or female hormone production. In mammals it is the female that is homogametic, and in birds it is the male. It has been proposed that sex chromosome asymmetries would result in the evolution of greater female sociality in mammals and male sociality in birds. In mammals, for example, a group of related females share more genetic material on the sex chromosomes with each other than a similar group of males do with each other. This is because brothers inherit their father's Y-chromosome, and one of their mother's X which could have come from either grand-father or grand-mother; sisters inherit one of their mother's X-chromosomes, and their father's X which must come from his grand-mother. This has been proposed as a reason for expecting females to be philopatric, but it may be over-ridden by the outcome of intra-genomic conflict.
HOMOLOGUE. One of a pair, or larger set, of chromosomes having the same overall genetic composition and sequence. In diploid organisms, each chromosome inherited from one parent is matched by an identical (except for mutational changes) chromosome - its homologue - from the other parent.
HOMOZYGOUS. Of a diploid organism having identical alleles of a given gene on both homologous chromosomes. An organism may be a homozygote with respect to one gene and, at the same time, a heterozygote with respect to another.
HYBRID. Cross between two varieties or species of plant or animal, differing in one or more genes.
HYBRID VIGOUR (HETEROSIS). The increase in size or rate of growth or fertility or survival (e.g. resistance to disease) associated with increased heterozygosity. It usually results from crosses between two genetically different, highly inbred lines.
HYBRID ZONE. An area of overlap between two species or sub-specific types where hybridisation takes place. For example the band of 24-170 km width of overlap between the hooded and carrion subspecies of crow.
IDEAL DESPOTIC DISTRIBUTION. A theoretical alternative to the IDEAL FREE DISTRIBUTION for explaining why individuals settle in less preferred habitats as population size increases. The ideal despotic distribution results from territory holders excluding newcomers from the preferred habitat and forcing them to settle in less preferred habitat where they experience reduced fitness (reproductive success, intake rates etc). Although all individuals within the same habitat therefore exhibit the same fitness, fitness will vary greatly among habitats (unlike in the IFD model). Further predictions of this model are a lower ratio of competitors to resources in good patches; and inverse spatial density dependence of prey mortality, because despotic exclusion is not compensated by increased intake for those remaining.
IDEAL FREE DISTRIBUTION. A theoretical model for the distribution of competitors between habitat patches [see Kacelnik, A. et al. 1992. TREE, 7:50; Tregenza, T. 1995. Adv. Ecol. Res., 26:253]. If all animals have equal competitive abilities (assumption 1) and are free to move anywhere without cost (assumption 2), and if resources occur as patches of different prey density (assumption 3), then initially the better quality patches will be preferred. But as the density of competitors on better patches rises, interference competition will increase, reducing intake rates to the same level as can be achieved on lower quality patches. It will then pay some animals to leave the better patches, leading to intake rates, or fitness, equalising across patches of different quality (prediction). This situation is termed the ideal free distribution (IFD). The model assumes a continuous input of resources (assumption 4; see Tregenza, T. 1994. Anim. Behav. 47:485 for common misapplications) with no depletion (assumption 5), and competitors who have full - ideal - knowledge of potential intake rates in all patches (assumption 6). Modifying this model to account for unequal competitors gives the IDEAL DESPOTIC DISTRIBUTION.
INBREEDING COEFFICIENT. The probability of obtaining homozygosity due to a zygote obtaining copies of the same gene from parents of common ancestry. Inbreeding increases the proportion of homozygotes above Hardy-Weinberg expectation, but does not change allele frequencies (although it can expose deleterious alleles to selection).
INBREEDING DEPRESSION. A loss of vigour amongst offspring occurring when closely related individuals mate, resulting from the expression of numbers of deleterious genes in a homozygous state and from a generally low level of heterozygosity.
INCLUSIVE FITNESS. The true measure of the fitness of an organism resides not only in its own success, but also in the influence it has on the fitness of other kin sharing some portion of its genotype. The idea is that helping relatives is a bit like helping yourself, because they share your genes. Inclusive fitness is defined as the animal's production of adult offspring, stripped of the average effect (across the population) of a single individual on others' reproductive success, and augmented by certain fractions of the quantities of the harm and benefit the individual itself causes to the fitnesses of its neighbours. The fractions in question are simply the COEFFICIENTS OF RELATEDNESS. [Abridged from Hamilton, W.D. 1964. J. theor. Biol., 7:1, amended following Creel, S. 1990. Proc. R. Soc. Lond. B, 241:229]. Individuals who attract more than an even share of help are of greater than average fitness, with regard to attracting help; those who attract less than an even share of help are of less than average fitness. For example, dwarf mongooses are cooperative breeders but have only one breeding pair per group which do not themselves provide help to others. If the average mongoose (including those that give no help at all) gives enough help in its lifetime to increase others' reproductive success by two offspring, then mongooses who receive sufficient help to raise their reproductive success by more than two offspring are more fit than average, with respect to attracting help. This is because mean help received equals mean help given when averaged across an entire population. The concept of inclusive fitness is most useful in modelling. It leads directly to HAMILTON'S RULE, however, which is more readily applicable to field data.
INTRAGENOMIC CONFLICT. The conflict of interests that is postulated to result from ultra-selfish genes acting to further their own evolutionary interests at a cost to the individual (genome) bearing them. Possibly a driving force behind the evolution of SEX, the SEXES and uniparental CYTOPLASMIC INHERITANCE. [See Hurst, L.D. 1992. Proc. R. Soc. Lond. B, 248:135].
INTRONS. Stretches of DNA sequences which divide the coding sequence of a gene into two or more parts termed exons. After the gene is transcribed into RNA, intron sequences are spliced out of the RNA sequence, a process called RNA splicing. First discovered in 1977, introns have been found in numerous eukaryote nuclear and organelle (chloroplasts and mitochondria) genes. Introns can be amplified by PCR, and they appear to have a similar rate of evolution to the gene, and higher than that of the axon (which is very conserved). Introns are therefore potentially of interest as GENETIC MARKERS. The base pair sequence differences should provide information directly about the rate of evolution, rather than producing banding patterns as in microsatellites and allozymes. [See Liu, X. 1991. BioEssays, 13:185.]
ISOGAMY. Fusion of gametes that are morphologically alike. This is an uncommon condition, found in some green algae, fungi and Protozoa.
ISOZYME (ISOENZYME). Different molecular forms of what is functionally the same enzyme. If two or more isozymes are coded for by different alleles at the same locus they are allozymes.
KARYOTYPE. The number and constitution of the chromosomes in a eukaryote cell. In multicellular organisms the somatic cells (all cells except the gametes) have the same karyotype
KIN SELECTION. The selection of genes due to one or more individuals favouring or disfavouring the survival and reproduction of relatives who possess the same genes by common descent. Coined by Maynard Smith (1964) to describe the process by which characteristics are favoured due to their beneficial effects on the survival of close relatives, including offspring (children and grand-children etc) and non-descendant kin (sibs and cousins etc). Possibly the most important mechanism for the evolution of ALTRUISM. See HAMILTON'S RULE for Hamilton's theory of kin selection.
KINSHIP. Possession of a common ancestor in the not too distant past. Kinship is measured precisely by the COEFFICIENT OF RELATEDNESS.
KOINOPHILIA. The tendency to choose mates with predominantly common phenotypic features. This form of MATE CHOICE may be an important factor in maintaining sexual reproduction, because maladapted traits tend to have low frequencies. In simulated competition between asexual and sexual reproduction with a mutation process creating mostly deleterious new alleles but a few beneficial, koinophilia allows sex to persist, whereas random mating results in bi-gender sex being rapidly replaced by asex. [Koeslag, P.D. & Koeslag, J.H. 1994. J. theor. Biol., 166:251]. Koinophilia can thus protect sex against invasion by parthenogens, despite sex paying the full two-fold cost of bi-gender reproduction: necessity for two individuals, one of which is a male, and even when males contribute nothing more than their gametes to their offspring. Since the cost of producing males is greatly discounted where males are involved in caring for their offspring, then the role of males is subject to evolutionary experimentation. This may explain the evolution of such a wide variety of functions that are fulfilled by males.
LETHAL-EQUIVALENT ALLELES. Alleles whose summed effect is that of lethality. For example, four alleles, each of which would be lethal 25% of the time (or to 25% of their bearers), are equivalent to one lethal allele. The average human carries about four lethal-equivalent alleles that are hidden as recessive alleles (estimated from inbreeding data).
LINKAGE. The presence on the same chromosome of two or more loci, usually recognised by the statistical tendency for alleles at linked loci to be inherited together. For example, if hair and eye colour are linked, offspring that inherit their mother's eye colour will also inherit her hair colour.
LINKAGE DISEQUILIBRIUM. The statistical tendency for alleles to occur together (e.g. fair-haired individuals tend to be blue-eyed).
LOCUS. The position of a gene in a chromosome.
MAINLAND-ISLAND MODELS. These ascribe the persistence of temporally varying habitat `islands' to dispersal entirely from a source `mainland.' Where mainlands are not themselves invulnerable to extinction by stochastic processes, the system can be represented by a METAPOPULATION: more or less isolated local populations not all concurrently extant, but interconnected by migration. See SOURCE-SINK models.
MAJOR HISTOCOMPATIBILITY COMPLEX (MHC). A large gene cluster that encodes various components of the immune system. Certain MHC genes can have many variant alleles; this produces an enormous diversity of antigens in a population, each individual possessing a unique set. The MHC also refers to the molecular pattern on the surface of each cell, which antibodies use to recognise the bodies own cells.
MARGINAL VALUE THEOREM. A hypothetical rule for OPTIMALITY MODELLING in situations where an individual alternates between exploiting patches and travelling between them. The policy for rate maximisation is to remain in the current patch as long as the expected pay-off per unit time in the next short period exceeds the expected pay-off per unit time in the environment as a whole. The MVT is therefore a useful tool for predicting individual behavioural decisions under different environmental conditions. A good correlation with field data suggests that the assumptions about the currency used by the animal, and the constraints on the animal's movements, are helpful in explaining its behaviour.
MATE CHOICE. Males can increase their reproductive success by mating with many females, because sperm is cheap. Females invest more in offspring than males, and can only increase their success by making eggs or young at a faster rate. In a 1:1 sex-ratio, females (and their investment potential) are therefore a scarce resource for which males compete; conversely females can afford to choose partners with genetic benefits, or with access to the resources which limit their investment potential. Thus it is commonly the males of a species that have adornments to enhance their competitive ability (e.g. antlers), and/or displays to enhance their attractiveness to females (e.g. bright coloration of polygynous male birds in contrast to cryptic coloration of the brooding female). The two opposing hypotheses for why females should prefer elaborate male displays or adornments are the FISHER and the HANDICAP hypotheses. Sometimes the general rule of high female investment is reversed and males are the main investors [Dale, S. & Slagsvold, T. 1994. Anim. Behav., 47:1197]. It is then the females which are competitive (e.g. polyandrous jacana females defend large territories in which several males incubate clutches), and the males which are choosy (e.g. humans in a monogamous system with male bread-winners). Females in a polygynous system may also compete among each other to attract male competition. For example, the prominent oestrus reddening and swelling around the perineum of Old World primate females is argued to be a sexually selected trait, reliably advertising an aspect of the female's condition. An individual benefits from a prominent swelling because it increases the males' willingness to compete for her, or because it draws their attention away from other females and towards herself: a form of spite. [See Pagel, M. 1994. Anim. Behav., 47:1333].
MATING SYSTEMS. The basic categories are MONOGAMY, POLYGYNY and POLYANDRY. The mating system adopted by an animal is determined by a number of factors, including (1) quantity of resources: in rich environments females can feed their young alone and are limited only by the number of eggs produced, favouring polygyny; in impoverished environments males gain more by sharing parental care with one female; (2) distribution of resources: patchy distributions result in males defending rich patches against rivals, favouring polygyny; (3) predation: monogamy may yield a higher return to the male if he helps to defend the brood, for example in colonial nesting seabirds where nest robbing is common; group living may enhance feeding efficiency or predator defence, resulting in polygyny particularly in ungulates and primates; (4) female availability in time: asynchronous females give a single male access to many females in succession, favouring polygyny; (5) requirements of the young: helpless young, e.g. of tree-nesting birds, require much parental care, favouring monogamy; precocious young, e.g. of many ground nesting birds, require less parental care, favouring polygyny.
MATRILINE. A social group of females comprising closely related individuals that persists for many generations (e.g. yellow-bellied marmot). Harems form when a single male attaches himself to one or more matriline. Matrilines are a common feature of mate-defence POLYGYNY (most mammals) in which DISPERSAL tends to be male-biased, and philopatry is favoured by females which are the limiting sex.
MEAN FITNESS. The mean value of the fitnesses of all the individuals in a population, calculated from the sum of the fitnesses of each genotype multiplied by the frequency of that genotype.
MEIOSIS. Cell division of a diploid cell to produce four haploid daughter cells. The process consists of two successive cell divisions with only one cycle of chromosome replication. It is a tenet of population genetics that meiotic segregation occurs independently of the fitness of the resulting zygote [but see Pomiankowski, A. & Hurst, L.D. 1993. Nature, 363:6428]. Moreover, the gametes produced after normal meiosis have a 50% probability of containing a given member of a pair of homologous chromosomes (the random assortment of chromosomes prescribed by traditional Mendelian genetics). But some instances occur where the segregation of chromosomes at meiosis is not fair and one of a particular pair of homologues is inherited more frequently than the other. This is described as MEIOTIC DRIVE.
MEIOTIC DRIVE. Any meiotic mechanism that results in the unequal recovery of the two types of gamete produced by a heterozygote. For example a gene which acts to promote its transmission by having a negative effect on the transmission of its homologue. Unfair segregation is most obvious where it is the sex chromosomes that are inherited in a biased manner, resulting in the offspring from a cross being predominantly of the same sex. [See Hurst, G.D.D. et al. 1992. TREE, 7:373].
MENDELIAN INHERITANCE. Non-blending inheritance by means of pairs of discrete hereditary factors (now identified with genes), one member of each pair coming from each parent. The main theoretical alternative is `blending inheritance.' In Mendelian inheritance genes may blend in their effects on a body, but they themselves do not blend, and they are passed on intact to future generations. (See also CYTOPLASMIC INHERITANCE).
METAPOPULATION. A theoretical concept describing an assemblage of spatially divided sub-populations whose units are interconnected by migration. One of its characteristic features is that not all its sub-populations are extant at the same time, reflecting a stochastic process of local extinctions and recolonisations which is dynamic over time. This results from the interplay of the local dynamics of births and deaths in individual sub-populations, with dispersal flow between patches of habitat. Levin's (1970) original concept of a metapopulation as a set of equally large patches all equally well connected to each other, has been developed with more realistic models, falling into three broad categories. Stepping-stone models: islands are distributed on a grid, and each is accessible to some neighbours but not all; give simultaneous solutions, useful for modelling coupled (e.g. predator-prey) systems. Reaction diffusion models: assume random dispersion (like molecules); reveal diffusion gradients between optimal and sub-optimal habitats, useful for modelling invasion of a new pest from known zero time. Incidence-function models: assume that extinction rates are a function of habitat patch area, and that colonisation rates are a function of isolation; this new category of models performs well in reproducing patterns observed in nature [see Hanski, I. 1994. TREE, 9:131]. The decreasing incidence of occupancy with increasing isolation and decreasing patch area parallels the predictions of the MacArthur-Wilson (1967) equilibrium theory for islands of varying size and isolation from a mainland source. Incidence-function models of metapopulation dynamics can be construed with a `mainland' (a very large patch from which all `islands' are colonised), but also without a mainland (migration occurring between many patches). These models are based on stochastic processes and assume the metapopulation is at a steady-state equilibrium, when rates of extinction and colonisation are in balance. Extinctions are often not due to stochastic processes, however, being associated with habitat degradation, and populations may not be at steady-state equilibrium if habitat has been lost recently [See Thomas, C.D. 1994. TREE, 9:300].
MICROEVOLUTION. A small amount of evolutionary change, consisting of minor alterations in gene proportions, chromosome structure, or chromosome numbers in a population. For example, (1) industrial melanism of peppered moths: the melanic form is under the control of single dominant gene that arises spontaneously by mutation from the colour gene; dark individuals are more frequent near industrial cities where light coloured lichens are rare, due to the selection force of insectivorous birds. (2) Balanced polymorphism in the sickle-cell trait: sickle-cell anaemia is caused by the abnormal haemoglobin S under the control of a single gene, lethal in the homozygous state; the trait survives because the heterozygous condition has superior fitness over both homozygotes: persons carrying haemoglobin S are more resistant to malignant tertian malaria. MACROEVOLUTION is a large amount of evolutionary change involving many elementary changes in gene proportions: the sum of many microevolutionary steps over a very large time-scale.
MICROSATELLITE MARKERS. Segments of DNA with tandem repeats of short-sequence motifs. They are numerous, highly variable and easy to score using the POLYMERASE CHAIN REACTION, making them excellent genetic markers for kinship studies. For example, a sequence for a wasp has a microsatellite consisting of ten A-A-T repeats. Other alleles at this locus have different numbers of repeats. When DNA at the microsatellite locus is amplified using PCR techniques and then electrophoresed through a gel to separate the alleles, each allele shows a characteristic three-band pattern. Alleles with fewer A-A-T repeats are smaller and migrate further down the gel. Microsatellites are unusual in that they mutate relatively frequently, generating new alleles that are thought to have little or no effect on an animal's fitness; if the members of a species mix and interbreed freely, then any new form of microsatellite will tend to spread homogeneously through the population. PCR primers can thus identify DNA motifs that evolve sufficiently rapidly to provide markers to characterise discrete populations. Character states are based on allele length, and therefore are not irreversible: individuals sharing alleles of the same length may do so by either relatedness or chance convergence (the mutation rate is estimated at 10-3 in mammals, so two populations more than a thousand generations apart cannot reliably be compared). An ideal system would be one in which new alleles are generated relatively frequently, but where the chances of back mutation are negligible (the `infinite alleles' model in which no alleles get mutated to twice). Minisatellite variable repeat (MVR) mapping may be a solution. Like DNA fingerprinting, this approach is based on minisatellites, but it maps the internal structure of a single locus (rather than measuring variations in length at one or several loci). [See Queller, D.C. et al. 1993. TREE, 8:285; Ashley, M.V. & Dow, B.D. 1994. In: Molecular Ecology and Evolution: Approaches and Applications. Berkhauser & Verlag, p:185; Wright, J.M. & Bentzen, P. 1994. Rev. Fish Biol. Fisheries, 4:384.]
MINIMUM VIABLE POPULATION (MVP). The minimum size of population that will ensure its persistence for a specified time within specified confidence limits. A common benchmark for MVP is 99% chance of persistence for 1000 years. MVP provides a focus for the study of ecological and genetic factors that influence the persistence of a threatened population. The principal approaches use either stochastic demographic models to determine how to minimise extinction probabilities, or genetic theory to determine how to maintain genetic variation (assuming that ability to evolve helps buffer populations against the unknown); both methods arrive at similar conclusions under panmictic conditions. [See Nunney, L. & Campbell, A. 1993. TREE, 8:234; also computer package described by Lacy, R.C. 1993. Wildl. Res., 20:45].
MITOCHONDRIA. Semi-autonomous organelles containing their own DNA and ribosomes and reproducing by binary fission, occurring in large numbers in the cytoplasm of eukaryotic cells. They are the major site of ATP production, and hence oxygen consumption in cells.
MITOSIS. Cell division in eucaryotes leading to the formation of two daughter cells each with a chromosome complement identical to that of the original cell.
MOLECULAR CLOCK. The idea that MOLECULAR EVOLUTION occurs at a constant rate, so that the degree of molecular difference between two species can be used as a measure of the time elapsed since they diverged. Its accuracy depends on the validity of the NEUTRALITY THEORY OF MOLECULAR EVOLUTION.
MOLECULAR EVOLUTION. Substitution of one amino acid for another in protein synthesis as a result of mutation of the genetic code. According to the NEUTRALITY THEORY OF MOLECULAR EVOLUTION, the variability at the molecular level which results from mutation is caused by random drift of the mutant genes rather than by selection.
MONOGAMY. A mating system in which an individual has one mating partner. About 90% of bird species are monogamous, but monogamy is very rare among other vertebrates. Its prevalence in birds is probably due to the demands of the young which require investment in incubation as eggs and in brooding as chicks. Both sexes therefore gain enhanced reproductive success by contributing to parental care. Monogamous females may engage in extra-pair copulations, or egg-dumping (laying eggs in another bird's nest), where reproductive success is limited by the number of eggs the female can incubate or chicks she can feed, and where this is lower than her reproductive potential. Adaptive advantages include: insurance against possible low fertility of a mate, and increasing genetic diversity through multiple paternity. Monogamous males may also have extra-pair copulations, to increase the number of their progeny, to increase the genetic diversity of their offspring, and to insure against having a sterile mate.
MULTIPLE INVASION. (allopatric speciation by). A few individuals of an original species breach an unchanging barrier to initiate a new and largely isolated population. Eventually the two populations evolve into separate species. If a second colonisation of the original species follows, the two daughter species will overlap without interbreeding.
MUTATION PRESSURE. Mutations are usually reversible, but if the rate in one direction is higher than in the other, then mutation pressure can progressively change the frequencies of mutating alleles.
MUTATION. Mutations are either point mutations, molecular substitutions of some nucleotide pairs for others in the DNA molecule, or chromosome aberrations, major structural changes encompassing hundreds or thousands of nucleotide pairs. Mutation rates are low, in the order of one in a million per generation, which are sufficient to create the variability needed to permit evolution to occur, but not great enough to drive evolution along by mutation pressure.
MUTUALISM. A type of symbiosis in which two or more individuals (or species) gain a net survival or reproductive benefit from co-operating or otherwise associating. For example, a lioness improves her capture success by hunting with another; she gains from the mutualism, and also receives kin-selection benefits if the other is related.
NATURAL SELECTION. The differential contribution of offspring to the next generation by individuals of different genetic types but belonging to the same population. This is the basic mechanism of evolution proposed by Charles Darwin and is generally regarded today as the main guiding force in evolution. It predicts `reproduction of the fittest' rather than survival of the fittest. It is the only one of the agents of evolution that specifically adapts populations to their immediate environment. In many animals it operates principally through differences in the reproductive success of individuals. Its result is always the same: some genotypes gain in the population at the expense of others. The selective force can act on the variability of a population in three different ways: a normal distribution of the population, which arises from many factors contributing jointly and independently to the trait under consideration, say size, can have both tails pulled in by stabilising selection, or one tail by directional selection, or be divided into two by disruptive selection. Natural selection is usually of overriding importance in MICROEVOLUTION.
NEO-DARWINISM. The current theory of the process of evolution, combining the Darwinian theory of evolution by natural selection with modern knowledge of genes and chromosomes to explain the source of the genetic variation upon which selection works.
NEUTRALITY THEORY OF MOLECULAR EVOLUTION (NEUTRAL-MUTATION THEORY) asserts that many genetic mutations are adaptively equivalent (effectively neutral), and do not affect significantly the fitness of the carrier. Thus they can become fixed in the genome at a random rate. Changes in their frequencies are due more to chance than to natural selection. The theory applies only to protein evolution and does not deny the role of natural selection in shaping morphological and behavioural attributes. (See MOLECULAR EVOLUTION).
OPTIMALITY MODELLING. A tool for the analysis of behavioural decisions in terms of their costs and benefits, on the assumption that natural selection tends to produce phenotypes that represent the best achievable balance of costs and benefits. An optimality model has three main components: a currency (e.g. maximising rate of energy intake, minimising risk of starvation); constraints (e.g. travel time, search time, handling time); the decision variable (e.g. whether to eat or reject food items, how much time to spend scanning, how much to invest in mate guarding). A good correlation of field data to the model suggests that the assumed constraints and currencies on which the model is based fit the real situation well. This technique is not suitable for individual behaviours that depend on what other members of the population are doing; here the correct approach is to model the EVOLUTIONARILY STABLE STRATEGY using GAME THEORY.
OUTBREEDING DEPRESSION. Reduction of fitness due to mating with unrelated individuals. Mating with relatives tends to conserve adaptive combinations of genes, but can also lead to INBREEDING DEPRESSION.
PANMICTIC UNIT. A local population in which mating is completely random. (See also DEME).
PHENOTYPE. The observable characteristics of an organism produced by the interaction of genes and environment. A gene may be said to have phenotypic expression in, say, eye colour.
PHYLOGENY. The evolutionary relationships within and between taxonomic levels, particularly the patterns of lines of descent, often branching, from one species to another.
PLASMID (PLASMAGENE). Cytoplasmic particle found in bacterial cells that carries one or more genes and that can replicate itself autonomously.
PLEIOTROPIC. A gene which affects more than one characteristic in the PHENOTYPE.
PLESIOMORPH. Primitive character state (see also APOMORPH).
POLYANDRY. A mating system in which a female maintains long-lasting and simultaneous relationships with several males. It is rare, contradicting the general proposition that males with the cheaper gametes are expected to be the polygamous sex. The system is exemplified by the jacana, where each female defends a large territory encompassing several smaller territories defended by males, each of which contains a nest. The female allocates all her reproductive effort to mating, egg-laying in all the nests, and defending her territory against other females. All the parental care of incubation and brooding is carried out by the males, which are smaller and subordinate to the females. The evolution of this system has been favoured by the scarcity of suitable breeding habitat, and heavy predation. Females enhance their reproductive success by laying large numbers of eggs, to exploit such suitable habitats as do exist and to offset losses to predation. Females are thus selected for large body size and resource allocation shifted towards egg production, at a cost to parental care which is delegated to the smaller, subordinated males. The female thus gains a higher reproductive success through her capacity to replace lost eggs than if she had to care for the young; the male also ensures a higher reproductive success for himself by caring for the young, than by leaving them prone to predation while seeking further matings. Males tolerate one another within a female's territory because they cannot be certain of paternity, being unable to distinguish between eggs in their clutch that they have fathered and those of other males as particular mating episodes are not linked to particular episodes of egg-laying.
POLYGENES. Genes with individually a very small effect on phenotype differences, systems of which are associated in producing quantitative variation of particular characters, for example stature.
POLYGYNY. A mating system in which a male mates with several females. Resource-based polygyny occurs where males control the females' access to resources that are essential to their reproductive effort, such as food or nest sites; where resources are patchily distributed, some males will successfully defend richer patches and attract many females. Female-defence polygyny occurs where males compete for females directly, with successful males holding a harem. Harems are facilitated if females show a predisposition to gather in groups, e.g. seals on pupping beaches. The mating success of males depends on their ability to compete with rival males, and to control the movements of females e.g. red deer stags chivvying hinds (hence the rarity of this mating system in birds which are highly mobile). Leks are a form of polygyny in which males control neither the resources nor females, but gather in dense clusters to display elaborate adornments (typically a ruff) and be chosen for mating by visiting females on the basis of plumage characteristics or display duration.
POLYMERASE CHAIN REACTION (PCR). Technique for amplifying a region of DNA using a pair of flanking primers and a thermostable polymerase. PCR primers are usually designed in order to amplify a specific locus, for example a dinucleotide simple sequence locus (MICROSATELLITE), based on a knowledge of the flanking nucleotide sequences.
POLYMORPHISM is the coexistence in the same population of two distinct hereditary types based on different alleles. For example the group of red cell polymorphisms affecting the haemoglobin molecule. These are maintained through superior malaria resistance in the heterozygous state despite being homozygous lethal. As well as such HETEROZYGOUS ADVANTAGE, polymorphism can also be maintained by FREQUENCY-DEPENDENT SELECTION in which the fitnesses of genotypes change according to their frequencies in the population (e.g. each genotype exploits slightly different resources). Parasite-maintained polymorphism may drive the evolution of SEXUAL REPRODUCTION and SEXUAL SELECTION by female MATE CHOICE (Hamilton & Zuk's version of the HANDICAP HYPOTHESIS). The idea relies on the facts that parasites, with shorter generation times and far greater fecundity than hosts, can evolve new genotypes faster than hosts, and that hosts are under continual selection for resistant genotypes. So genetic diversity of hosts may be selectively maintained by parasite genetic diversity. [See Gulland, F.M.D. 1993. Proc. R. Soc. Lond. B., 254:7].
POLYPHYLETIC. A group of species classified together is polyphyletic when some of its members have had quite distinct evolutionary histories, not being descended from a common ancestor which was also a member of the group; so that, if the classification is to correspond with phylogeny, the group should be broken up into two or more distinct groups.
POLYTHETIC. System in which membership of taxon is based on possession of a large number of common characters.
POLYTOPIC. (of a taxon). Occurring in two or more separate areas.
POSITIVE ASSORTATIVE MATING. The `Wallace Effect': mating preferentially with individuals of ones own population or putative species. Also known as `reinforcement' and its action is to reduce the production of hybrids.
PREADAPTATION. A phenomenon whereby some structures in ancestors are fortuitously suited for transformation to a new adaptation in descendants. Preadaptation helps to explain the appearance of fundamentally new designs, such as the feathers on bird wings (what good would be an embryonic wing to flight?) or the vertebrate jaw.
PROKARYOTES. Organisms in which the genetic material is not enclosed in a cell nucleus (e.g. bacteria). EUKARYOTES are thought to have evolved as symbiotic associations of prokaryotes.
RADIATION. An episode of significantly sustained excess of cladogenesis over extinction in a clade of organisms. The proliferation of species into different niches of a community is called adaptive radiation. The similarity acquired by species that come to occupy approximately the same ecological position but in different communities is called CONVERGENCE. The marsupials of Australia have undergone adaptive radiation during the past 70 million years, filling most of the niches open to land-dwelling mammals. In other continents equivalent radiation was achieved by placental mammals, resulting in an astonishing degree of convergence (e.g. placental versus marsupial wolves, moles and flying squirrels). In addition, unique forms have been produced on both sides, e.g. kangaroos are the large herbivores of Australia. Another example of adaptive radiation by speciation is the 22 species of Hawaiian honeycreepers, which probably originated from a single species of finch-like bird that colonised the islands and later radiated by means of MULTIPLE INVASION. Speciation occurs easily among the expanding populations, because the water gaps separating the islands are powerful barriers to gene flow between populations inhabiting different islands. As the species diverged from each other, they tended to expand quickly into the unfilled ecological positions that were so abundantly available on these remote islands, resulting in broad variation among a relatively small number of species.
RECOMBINANT DNA TECHNOLOGY (GENETIC ENGINEERING). Manipulation of DNA using restriction enzymes which can split the DNA molecule and then join vector with foreign DNA to form a hybrid molecule of non-homologous DNA called recombinant DNA. This technique of GENE CLONING by-passes all the biological restraints to genetic exchange and mixing, and even permits the combination of genes from widely different species.
RED QUEEN'S HYPOTHESIS. No matter how well members of a species are adapted at any one time, their environment can be said to be constantly deteriorating with respect to their adaptations. Adaptive responses to one selective pressure may be deleterious to adaptations to other, less immediate pressures, so that mean fitness declines whenever the balance of pressures changes. The balance of selective pressures is constantly varying, for example through the continuing adaptive responses of different species to each other, so an advantage gained by one may correspond to a worsening of the environment experienced by another. Mean fitness in a species therefore fluctuates randomly with time, and the risk of extinction is constant for all species within an ADAPTIVE ZONE. First proposed by Van Valen (1973) to explain why probabilities of species extinction were not observed to change with taxon duration (unlike individuals which degenerate towards extinction at an increasing rate with age). Red Queen dynamics are postulated to underpin SEXUAL REPRODUCTION and SEXUAL SELECTION through parasite-maintained POLYMORPHISM.
REPRODUCTIVE EFFORT. The total resources of time and energy used by an animal in reproduction, partitioned into parental effort (provisioning and rearing offspring) and mating effort (acquiring mates).
REPRODUCTIVE SUCCESS. can be measured by counting the number of offspring produced by an individual, often suitable in field studies where mean numbers can easily be compared between groups of interest). An equivalent measure is INCLUSIVE FITNESS, usually more suitable for modelling when the hypothetical trait in question is defined in terms of differences in numbers of offspring. Inclusive fitness is relevant in field studies when the logic of an animal's decision rests with differences in reproductive success, in terms of numbers of offspring, and when we can measure those differences through experiment or through natural variation. The factors affecting reproductive success commonly differ between the sexes: see SEXUAL SELECTION.
RIBOSOMES. Granules composed of RNA and protein found in large numbers in all types of cells, where they synthesise protein.
RNA (RIBONUCLEIC ACID). Nucleic acid characterised by the presence of D-ribose and the pyrimidine base uracil. It occurs in three principal forms, as messenger-RNA, ribosomal-RNA, and transfer-RNA, all of which participate in protein synthesis.
SECONDARY SEXUAL CHARACTER. Characteristic of animals that differs between the two sexes, other than the gonads, ducts and glands that convey the gametes (e.g. mammary glands, external genitalia, antlers, peacock's tail).
SELECTION DIFFERENTIAL. The amount by which the mean value of a character among those individuals that survive to reproduce differs from the mean value for the whole population.
SELECTION PRESSURE. Any feature of the environment that results in natural selection; for example, food shortage, the activity of a predator, or competition from other members of the same sex for a mate, can cause individuals of different genetic types to survive to different average ages, to reproduce at different rates, or both.
SEX (MIXIS). The exchange of genetic material between individuals. Gender and reproduction are incidental, though widespread, associations with sex.
SEX. (evolution of). The genetic diversity associated with sexual reproduction clearly enhances the survival of species, but it is less easy to understand as an adaptation at the individual level. A conceptual framework provided by modelling INTRAGENOMIC CONFLICT suggests how sex can be seen as an adaptation with fitness benefits for selfish genes. Sex may initially have evolved as a means by which an ultra-selfish gene could increase its transmission frequency at a cost to the host genome. A gene without such a means of transfer is stuck in the vertical lineage that descends from the original parent cell, but a gene with horizontal transmission capability has the potential to infect numerous other lineages. For example, plasmids that inhabit the mitochondria of a slime mould appear to force the fusion of two isogamous gametes. If one set of mitochondria has the plasmid but the other does not, then the mitochondria fuse, recombine, and split apart again, after which the plasmid is found in all the subsequent mitochondrial products. Fusion is not witnessed in the absence of the plasmid. If the plasmid is not suppressed by the host genome, then the initial cost of establishing the sexual process may be overcome; once established, however, the host genome can take control of the parasitic selfish gene and adapt it to its own advantage by hooking up host DNA to the plasmid. After this evolution of cytoplasmic fusion of gametes, a new conflict arises: between maternally and paternally derived cytoplasmic genes for control of the zygote. If cytoplasmic genes from one parent attempt to annihilate their competitors from the other parent, this sort of aggressive behaviour might lead to the evolution of SEXES. [See Hurst, L.D. 1992. Proc. R. Soc. Lond. B, 248:135].
SEXES. (evolution of). Most organisms have two sexes, some have none (e.g. most ciliates), but the incidence of more than two sexes is very rare (some acellular slime moulds). In the most rudimentary differentiation into mating types, the gametes are isogamous, suggesting the view that sperm evolved to prevent the mixing of cytoplasmic genes from different parents. The fundamental asymmetry of the sexes therefore evolved as a means to minimise the damage caused by conflict between cytoplasmic genes in the zygote. For the common case, the male sex is that which resigns attempts to contribute cytoplasmic genes to the next generation (see CYTOPLASMIC INHERITANCE). Species with more than two sexes may also have uniparental inheritance of cytoplasmic genes; they are nevertheless rare and could be afflicted by deleterious cytoplasmic genes which attempt to pervert normal cytoplasmic genes. This is because a system in which the asymmetry of sex is established only after zygote formation is inherently more vulnerable to selfish, zygote-injurious cytoplasmic genes than one in which the asymmetry is established before fusion (as for two sexes). For example the system will be vulnerable to selfish cytoplasmic genes which attempt to prevent uniparental inheritance of cytoplasmic genes. One such gene is known to be carried by a plasmid in the mitochondria of a slime mould, which forces fusion of mitochondria at zygote formation. After splitting apart, all the mitochondria have this plasmid. If this effect is costly for the nuclear genes, then a gamete which preferred to avoid this problem by destroying its own cytoplasmic genes and mating with those gametes which are prepared to let their cytoplasmic genes be transmitted (proto-females) could spread if the advantages outweighed the cost: hence the collapse of multiple sex systems to binary types. [See Hurst, L.D. & Hamilton, W.D. 1992. Proc. R. Soc. Lond. B., 247:189].
SEX CHROMOSOME. In organisms with a chromosomal mechanism of sex determination, one of the chromosomes involved in sex determination. One sex chromosome, the X-chromosome, is present in two copies in one sex and only one copy in the other sex. The autosomes, as opposed to the sex chromosomes, are present in two copies in both sexes. In many organisms, there is a second sex chromosome, the Y-chromosome, that is found in only one sex - the sex having only one copy of the X. Sex-linked genes are those carried on the sex chromosomes.
SEXUAL REPRODUCTION. Reproduction involving the fusion of haploid nuclei, usually gametes, which result from MEIOSIS. Sexual reproduction creates diversity but does not force changes in gene frequencies. It is therefore the agent of adaptability but not of evolution. The population uses sexual reproduction to increase the diversity it needs to evolve, but sexual reproduction alone does not cause the population to evolve. Instead, the essential changes in gene frequencies are caused by four evolutionary agents: MUTATION PRESSURE, GENETIC DRIFT, GENE FLOW, NATURAL SELECTION.
SEXUAL SELECTION. Selection for traits which are solely concerned with increasing mating success, either by favouring the ability of one sex (usually males) to compete directly with one another for fertilisations, for example by fighting (intra-sexual selection), or by favouring traits in one sex which attract the other (inter-sexual selection: see MATE CHOICE). Where natural selection operates through differences in the reproductive success of individuals, the factors affecting RS commonly differ between the sexes: since the energetic costs of reproduction are usually higher in females than males, in a population with 1:1 sex-ratio (see FISHER'S SEX RATIO THEORY) it is access to females and their investment capability which usually limits the reproductive success of males, while access to resources usually limits that of females. Where access to resources depends mainly on success in indirect competition but access to females depends principally on direct competition, traits promoting fighting ability (e.g. antlers) will be more strongly favoured in males than females, while those promoting feeding efficiency will be more strongly favoured in females. It is usually more useful to attempt to relate traits that are more developed in one sex to differences in the selection pressures operating on males and females than to try to distinguish between differences that are a consequence of natural selection and those that have been produced by sexual selection. For example, if body size confers advantage on males, and large mothers produce large sons, competition between males may select for increased body size in females. [See Clutton-Brock, T.H. 1982. Red Deer: Behaviour and Ecology of two Sexes. Edinburgh University Press].
SOURCE-SINK MODELS. These models ascribe the persistence of qualitatively inferior local populations (sinks) in spatially varying habitat to dispersal entirely from a source population in which reproduction exceeds mortality at equilibrium population size. Evidence for source-sink systems is hard to obtain: viable populations may appear as sinks because dispersal into them depresses fecundity or increases mortality as a result of density-dependence; local recruitment then appears insufficient to balance local mortality. [See Watkinson, A.R. & Sutherland, W.J. 1995. J. Anim. Ecol., 64:126.]
SPECIATION can result from two different mechanisms: allopatric (geographic) speciation, by simple FISSION or by MULTIPLE INVASION; sympatric speciation, by polyploidy (accidental doubling in number of chromosomes in some offspring which then interbreed among themselves) or by disruptive selection (new selective force splits a previously freely interbreeding population by removing intermediate individuals). Geographic speciation is the most frequent in animals and plants; polyploidy is insignificant in animals, but responsible for the origin of about half of the extant higher plant species; disruptive selection has not been demonstrated.
SPERM COMPETITION. Competition between sperm from different males to fertilise the ova of a female. This is a widespread phenomenon among animals with external fertilisation (e.g. fish), but also among mammals in species where the female stores sperm between mating and fertilisation and can mate with more than one male during this period. A male's probability of paternity can be influenced by factors such as the delay between copulations of competing males, the number of ejaculations attained by each male, and the timing of copulation relative to female ovulation. EVOLUTIONARILY STABLE STRATEGIES can be modelled under conditions of sperm competition between unequal males to predict that small males should invest more heavily in sperm than dominant males, which have access to females at the optimal time for achieving fertilisation. The males of some species have adaptations to improve their certainty of paternity: male rats leave a gelatinous plug in the female's reproductive tract after mating; male damselflies remain attached to the female between mating and egg-laying, with a consequent reduction in the number of females that can be mated; house sparrow pairs mate up to 40 times per day during the female's egg-laying period.
TENSION ZONE. Where two adjacent closely related species (often sister species) have expanded to meet and produce a HYBRID ZONE which is maintained by selection against hybrids. The tension is between selection against the hybrids, causing the zone to narrow, and the lifetime movement (vagility) of the hybrids themselves causing the zone to expand with introgression each side into the parent populations. The former leads to speciation, the latter to merging of the two species.
TERRITORIAL BEHAVIOUR. Behaviour that leads to individual animals or groups being spaced further apart than would be expected from a random occupation of suitable habitats. Territorial behaviour functions to ensure that individuals have sufficient food, shelter or mates to rear their young. The effect of territoriality on the genetic composition of the population, however, is to restrict the proportion of the total population in any one year that breeds and thus contributes genes to the next generation.
TRANSGENIC. A transgenic organism has a genome that incorporates and expresses genes from another species. Such organisms are created by genetic engineering, using suitable cloning vectors (e.g. retroviruses) to insert the desired foreign gene into the fertilised egg or early embryo of the host. Transgenic sheep have been engineered to produce therapeutic proteins in their milk; transgenic crops can be engineered for improved disease resistance.
VICARIANCE. The initiation of SPECIATION and thus CLADOGENESIS by the origin ('vicariance event') of a barrier dividing the range of the parent species. It can be expected to affect a whole number of not necessarily closely related species in the same way.
VIRULENCE. The disease producing ability of a micro-organism. Natural selection between hosts tends to favour lower virulence of a pathogen. If dispersal of the pathogen depends on the host's survival and mobility, any damage to the host is harmful to the pathogen. Thus diseases spread by personal contact, such as the cold virus, tend not to be highly virulent. Natural selection within a host, however, tends to favour extreme virulence of a pathogen. It exploits its host to maximise the current rate of dispersal of new individuals to new hosts. It may kill its host quickly but while the host lives it does better than competing pathogens. Thus the malaria agent Plasmodium gets no benefit from the host's feeling well; in fact a prostrate host is more vulnerable to mosquitoes.
VIRUS. Type of non-cellular organism which has no metabolism of its own. It consists mainly or solely of nucleic acid genome (RNA or DNA) enclosed by protein. To replicate itself, a virus must infect a cell of a suitable host organism where it redirects the host-cell metabolism to manufacture more virus particles. Viruses can reproduce sexually, but in a way different from eukaryotes. Whenever two or more viruses co-infect the same host cell, hybrid progeny can be produced through genetic exchange between co-infecting parent viruses [see Chao, L. 1992. TREE, 7:147].
WILD TYPE. Used by laboratory geneticists to refer to the `normal' allele at the same locus as a conspicuous mutant, or to the normal organism. In other words, the phenotype of a particular organism when first seen in nature.
ZYGOTE. The fertilised ovum of an animal or plant formed from the fusion of male and female gametes.
Source books
Alberts, B. et al. 1983. Molecular Biology of the Cell. Garland Publishing, New York.
Allaby, M. (ed.) 1985. The Oxford Dictionary of Natural History. Oxford University Press, Oxford.
Begon, M., Harper, J.L. & Townsend, C.R. 1990. Ecology. Individuals, Populations and Communities. Blackwell Scientific, Oxford.
Dawkins, R. 1982. The Extended Phenotype. Freeman, Oxford.
Krebs, J.R. & Davies, N.B. (eds). 1991. Behavioural Ecology: An Evolutionary Approach, 3rd Ed. Blackwell Scientific, Oxford.
Krebs, J.R. & Davies, N.B. 1993. An Introduction to Behavioural Ecology, 3rd Ed. Blackwell Scientific, Oxford.
Martin, E. 1990. A Concise Dictionary of Biology. Oxford University Press, Oxford.
Nesse, R.M. & Williams, G.C. 1995. Evolution and Healing. Weidenfeld & Nicolson, London.
Skelton, P. (ed.) 1993. Evolution. A Biological and Palaeontological Approach. Addison-Wesley, Wokingham (The Open University).
Tamarin, R.H. 1993. Principles of Genetics, 4th ed. WCB, Oxford.
Wilson, E.O. et al. 1978. Life on Earth, 2nd Ed. Sinauer Associates, Sunderland, Mass.
See also the Lexicon of
Reproductive Modes, and the Lexicon of
Statistical Modelling. For further information, try the Biotech Life
Science Dictionary.